
SEV Core Site plant phenology monitoring and mast fruiting
Objective:
Phenology, the timing of recurring life cycle events, is an important metric for understanding seasonal variation within biological systems. Because the timing of life cycle events is often sensitive to environmental conditions, climate change can have strong effects on phenology, with critical consequences for species interactions and ecosystem processes. Reproduction, a key component of the plant life cycle, is necessary to predict future plant and wildlife population abundances. Accurate prediction is particularly challenging for plants with highly variable fruit production, known as mast fruiting. We document the phenology of plants and mast fruit production in five dryland ecosystem types to monitor changes in seasonal patterns over time and in conjunction with our drying and increasingly more variable climate.
Novelty:
Our datasets are unique in their longevity and robustness because they can be paired with SEV LTER small mammal, grasshopper, bee, and plant biomass datasets to study spatial and temporal trends in consumers, pollinators, and plant communities. The tree mast production study is the longest, continuously ongoing study on mast fruiting to our knowledge, and tracks >100 individual trees from three foundation tree species.
Design:
Plant phenology data are collected once per month from February through October along four replicate 200 m transects in each of three core ecosystem types: Chihuahuan Desert grassland, dominated by black grama grass; Plains grassland, dominated by blue grama grass; and Chihuahuan Desert shrubland, dominated by creosotebush, all within the Sevilleta National Wildlife Refuge. On each transect, we record the foliage and reproduction status of the first 10 individuals of each plant species encountered. Foliage is an assessment of the plant’s leaves (e.g., presence of new green leaves, old leaves, brown leaves) while reproduction examines flower and fruit stages (presence of buds, flowers, fruits). We measure annual mast fruit production in August at five sites within the Sevilleta National Wildlife Refuge. For each tree species, we use a specially customized protocol to estimate fruit production. piñon pine (Pinus edulis) estimates are made using the number of cones per mature tree. Shrub oak (Quercus turbinellla) estimates utilize the number of acorns per 0.1 m2 of canopy surface area. One-seed juniper (Juniperus monosperma) estimates use the number of berries per 0.5 m2 frame, assessed at four aspects per individual plant, then summed for each tree in the one ha plot.










The tree mast production study is the longest, continuously ongoing study on mast fruiting to our knowledge, and tracks >100 individual trees from three foundation tree species.
Responses:
The plant phenology dataset provides population-level estimates of the proportion of individuals in each foliage and reproduction stage during a given month for each plant species x transect combination, and data have been collected continuously since 2000. The mast fruiting dataset gives annual fruit production for three tree species: piñon pine (Pinus edulis), Shrub oak (Quercus turbinellla), and One-seed juniper (Juniperus monosperma), and data have been collected since 1997.
Supporting Documents:
Plant phenology dataset:
Core Site Phenology Study from the Chihuahaun Desert Grassland and Shrubland at the Sevilleta National Wildlife Refuge, New Mexico. https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=137
Mast fruiting dataset:
Tree Mast Production in Pinyon-Juniper-Oak Forests at the Sevilleta National Wildlife Refuge, New Mexico (1997- present). https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=204

SEV Plant Traits Monitoring
Objective:
Traits and diversity are key to evaluating why some ecosystems and species are more sensitive to environmental change than others. Traits are essential to generalizations beyond the dynamics of specific species. Traits also help us to understand the biophysical feedbacks between plants and the abiotic environment. Therefore, we have assembled existing and newly collected data into the first SEV LTER Trait Database. Monitoring trait change is critical to effective prediction because altered trait distributions will shape how future populations and communities respond to amplified environmental variability. We are currently evaluating whether long-term increases in climate mean and variance will shift trait distributions from acquisitive toward more conservative strategies.
Novelty:
Historically, LTER sites, including ours, focused on patterns and process from a largely species- or community-centric rather than traits-based perspective. Because traits add novel information on species’ functions in ecosystems they can increase the information yield from analyses of long-term dynamics, their drivers, and their consequences.
Design:
We follow established protocols for standardized plant trait data (Perez-Harguindeguy et al. 2013) to sample at least 10 individuals per species across diverse dryland biomes of the SEV LTER program.




Plant traits measured at SEV core sites.
Responses:
We focus on traits important to the performance and persistence of primary producers in drylands. Leaf traits that mediate water balance, tolerance to drought, and herbivory include specific leaf area, water use efficiency, and leaf [C]:[N] ratio. Belowground traits inform on plant responses to precipitation and include rooting depth and specific root length. Other plant traits such as height, lifespan, seed mass, and dormancy affect plant responses to climate and species interactions. We have also added field-collected spectral reflectance data for a subset of focal plant species.
Supporting Documents:
Field-Collected Spectral Reflectance of Dominant Vegetation at the Sevilleta National Wildlife Refuge. https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=328

Burning Experiment (BurnX)
Objective:
Disturbance from fire can alter the abundance and distribution of shrubs, grasses, and forbs in dryland ecosystems. In general, fire is expected to increase grass and forb production while hindering shrub encroachment in drylands. Therefore, prescribed fires are a common management tool for maintaining grassland habitats in the southwestern USA. However, black grama (Bouteloua eriopoda), a dominant species in Chihuahuan Desert grassland, can be highly susceptible to fire, potentially resulting in death followed by slow recovery rates. In contrast, blue grama (B. gracilis), the dominant of the short grass steppe and Plains grasslands may be more resistant to repeated fires. A 2003 prescribed fire in a mixed grassland of blue and black grama the Sevilleta National Wildlife Refuge in central New Mexico offered the opportunity to study the long-term effects of fire on both of the dominant species and a cohort of other grass, shrub and forb species in this region. This study was conducted on a site that had a previous prescribed burn in 1993, allowing us to incorporate previous fire history into the study design.
Novelty:
There are few long-term records of the impact of fire in semi-arid and arid, desert grasslands. Most fire studies in drylands have limited temporal scope. Accordingly, how fire is perceived and used in dryland ecosystems of the southwestern USA often relies on information derived from mesic ecosystems, such as short grass steppe, wet prairies, or even woodlands. Here, we have tracked occupancy by individual grass patches, post-fire accumulation of plant biomass, and plant species cover and diversity dynamics at decadal scales that provide insight into important disturbances in drylands. Our results inform land managers on appropriate time scales for managing fire in dryland landscapes of the desert southwest.
Design:
The BurnX study was established in a mixed black and blue grama-dominated grassland in a randomized block design consisting of 16, 9-ha blocks (300 m x 300 m) arranged in a 4×4 grid. Within each block are 12 random permanently marked subplots, of which 6 are used for the post-2003 fire measurements. Bulk biomass by year is measured in the fall by clipping two, 25 x 50 cm quadrats extending out from the subplot corner stakes in new locations in each sampling period. Occupancy by species is repeatedly measured in four, gridded 30 x 30 cm quadrats at the corner stakes of each subplot. In addition, each block contains a standard Sevilleta mammal trapping web which consists of 12 transects radiating from a central point. Each transect contains 12 numbered stakes for a total of 144 potential sample locations per web. Vegetation species composition and cover is measured in permanently located 50 cm x 50 cm quadrats at four evenly spaced points along each web transect (48 permanent quadrats per replicate) with only the first four even numbered stakes from the end of the transect used.




Yucca burning, fire monitor and burning landscape. Field work in progress.
Responses:
Before and after the 2003 fire, above ground plant biomass accumulation and species composition were monitored from 2003 to 2018. Vegetation cover and biomass by species is estimated visually in the spring and fall each year on the web transects. Spring measurements are taken in May when spring annuals have fully matured; and fall measurements are taken in September or October when summer annuals have fully matured and perennial species have reached their peak biomass for the year before the frost. Subplot measurements of biomass and occupancy are taken only in the fall.
Discoveries. Following prescribed fire, there were fewer black grama individual grass clumps and less aboveground grass biomass and cover in burned areas than unburned plots. The recovery rate of black grama grass was slow during the 15 years following the fire. Other grasses, particularly blue grama, experienced limited to no adverse effects from fire. Similarly, impacts of fire on shrubs and forbs were limited, and overall plant species diversity was unaltered.
Supporting Documents:
2003 Prescribed Burn Effect on Chihuahuan Desert Grasses and Shrubs at the Sevilleta National Wildlife Refuge, New Mexico: Grass Recovery Study (2003-2018). https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=148
Burn Exclosure Rodent Population Study at the Sevilleta National Wildlife Refuge, New Mexico (1991-1993). https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=15

Cactus Demography, Population Biology, and Plant-Animal Interactions
Objective:
The population dynamics of plants are driven by demographic rates of growth, survival, reproduction, and recruitment, and the influence of biotic and abiotic factors on those demographic processes. This study is designed to understand plant population dynamics in response to climate fluctuations, density-dependent feedbacks, and ecological interactions with animal mutualists and antagonists. Long-term demographic data provide the foundation for building models that allow us to forecast or “back-cast” population responses to past, present, and future environmental change. The focal species of this study is the tree cholla cactus, Cylindriopuntia imbricata.
Novelty:
Initiated in 2003, this study has become one of the longest longitudinal studies of plant demography and population dynamics. The long time series allows us to connect inter-annual demographic variation to weather data, to detect trends in plant susceptibility to insect herbivores, and to observe shifts in associations with defensive mutualists (extrafloral nectar-feeding ants).
Design:
Longitudinal demographic data are collected annually in late May from tagged plants within eight 30m x 30m plots in the foothills of the Los Piños mountains. Plots are searched for new recruits each year, and these are added to the census.




Cylindriopuntia imbricata, the Tree Cholla Cactus, is the focal species of this study.
Responses:
We record plant survival, size, reproductive output, number and identities of cactus-feeding insects, number and identities of ant mutualists. Numbers of new recruits in each plot are recorded each year. Individual plants are GPS’d to enable spatial analyses.
Supporting Documents:
Long-term study of tree cholla demography in the Los Pinos mountains, Sevilleta National Wildlife Refuge. https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=323

Drought Network (DroughtNet) experiment
Objective:
The Drought Network is a series of globally distributed drought experiments with a standardized protocol which provides the capacity to understand drought sensitivity patterns across biomes and climate gradients. Three Sevilleta sites were established in 2018 in Chihuahuan Desert grassland (black grama-dominated), Plains grassland (blue grama-dominated), and an ecotonal site. Together, these sites contribute data to the global network, but also provide the opportunity to study short-term drought effects across Sevilleta grasslands. The three Sevilleta sites allow us to compare drought responses between dominant grass species, black grama and blue grama, both separately and while in direct competition.
Novelty:
The Sevilleta sites were installed in coordination with four other sites in the Sonoran and Mojave deserts, which when combined, make up one of the driest sets of coordinated drought experiments in the world. By studying plant, biocrust, microbial and fungal responses to drought across three North American deserts, we can begin to understand the mechanisms by which state change occurs in response to drought and predict future changes in communities due to climate variability and associated extreme climate events.
Design:
Each site includes fourteen plots (2.5 m x 2.5 m): seven control plots and seven treatment plots with structures that reduce ambient rainfall year-round by 66%. A 1 m x 1 m permanent quadrat located in the center of each plot is used for plant monitoring, and other processes are measured around the perimeter of the quadrat. We use existing control plots at EDGE and WENNDEx experiments for control plots because the DroughtNet infrastructure is co-located with these experiments. The drought treatment will be applied for four years (until 2022), at which time the drought shelter infrastructure will be removed and post-drought recovery monitoring will begin.




DroughtNet sites
Responses:
Each spring and fall, we measure plant cover and estimate biomass by species using standard SEV LTER protocols. Seasonal biocrust monitoring began in 2019, and collections for biocrust and fungal communities, microbial biomass, and plant stable isotopes occurred in 2020 after two years of drought treatment.
Supporting Documents:
Drought Impact on Desert Ecosystems, Drought Network precipitation manipulation experiment in desert grasslands. https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=336

Fire Seasonality Experiment
Objective:
To determine whether seasonality of the timing of prescribed fire matters for management fires in Sevilleta grasslands.
Novelty:
The regional climate consists of two precipitation periods, winter/spring rains and the summer monsoon separated by a ~2 month hyperarid period. Managers assumed that natural fires occurred primarily during the hyper-arid period prior to the monsoon, when fuel is available and dry lightning strikes often occur. As a consequence, management burns were scheduled for this period, but regional wildfires often prevented management burns from taking place. The purpose of the ongoing Fire Seasonality experiment is to determine if more flexible burning schedules allow for management burns to occur when local fire crews are less occupied fighting regional wildfires.
Design:
The study consists of 20 plots each of which is 20 m x 40 m. Four treatments include unburned or management burns applied in mid-March, late-June, or late-November, with n = 5 replicates of each treatment. Experimental fires are carried out by the local Fish and Wildlife Service fire crew. Thus far, each plot has been burned twice: 1) November 2007, March 2008 and June 2008, and 2) November 2016, March 2017 and June 2017.



Burnplots, controlled burn video, and site map
Responses:
Each replicate plot has 10 permanently located 1 m x 1 m quadrats for estimating plant species composition with visual estimates of cover by species. Vegetation sampling occurs in spring and fall annually. In fall 2018, all plots were also sampled with high resolution (5 cm) drone imagery.
Supporting Documents:
Effects of Fire Seasonality on Chihuahuan Desert Grasslands at the Sevilleta National Wildlife Refuge, New Mexico (2007-2020). https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=230
2003 Prescribed Burn Effect on Chihuahuan Desert Grasses and Shrubs at the Sevilleta National Wildlife Refuge, New Mexico: Species Composition Study (2004-2018). https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=166
2003 Prescribed Burn Effect on Chihuahuan Desert Grasses and Shrubs at the Sevilleta National Wildlife Refuge, New Mexico: Shrub Recovery Study (2003-2009). https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=164
Ladwig, L.M., S.L. Collins, P. Ford and L.B. White. 2014. Chihuahuan Desert grassland responds similarly to fall, spring, and summer fires during prolonged drought. Rangeland Ecology and Management 67: 621-628. https://doi.org/10.2111/REM-D-13-00133.1

Foundation Plant Species Loss Experiment
Objective:
This experiment evaluates the responses of plant species composition, net primary production (NPP), and ecosystem processes to the removal of dominant, foundation plant species. The experiment was initiated by Deb Peters (NMSU, JRN LTER) in 1995 to address questions about competitive interactions between foundation species and has since been maintained annually by the SEV LTER Field Crew. In addition to the original vision, we repurposed this experiment to address: How will changes in climate variance and mean affect ecosystem-specific biogeochemical processes and alter biophysical feedbacks?
Novelty:
Changes in climate mean and variance could alter the strength of biophysical feedbacks between plants and the abiotic environment in ways that accelerate or inhibit ecosystem transitions, dramatically altering predictions on future states. It remains unclear whether or when such changes will occur in drylands experiencing greater inter-annual climate variance. Plant species removals provide a powerful tool to assess feedbacks, by comparing process rates in the presence versus absence of a foundation plant species. These experiments provide a window on how biogeochemical and biophysical processes, as well as other community members, respond to the loss of foundation species. This experiment also enables the collection of new plant demographic data in the presence/absence of competitors to inform our predictions of the trajectories of ecosystem transitions, using our WAVE model (see Models Overview). Finally, plant removals have led to new tests of how biodiversity-function relationships change when foundation plant species are lost from ecosystems.
Design:
We selected five sites that were dominated by either blue grama (site 1), blue and black grama (site 2), black grama (site 3), black grama and creosote (site 4), or creosote (site 5). A sixth site was later added in the blue grama community along the foothills of the Los Pinos Mountains (site 6) near the Blue Grama Core Site (see Core Site Overview). At sites 1, 3, 5, and 6, five 3 m x 4 m plots had all plants of the dominant species removed initially and then allowed to recolonize; five 3 m x 4 m plots were controls. At site 2, 5 plots have blue grama removed annually, 5 plots have black grama removed annually, and 5 plots are controls. At site 4, 5 plots have black grama removed annually, 5 plots have creosote removed annually, and 5 plots are controls. Initial cover prior to removal was estimated by species for each plot. Grass was removed using a shovel to collect above-ground biomass and crowns just below the soil surface. Shrubs were removed using large clippers to collect above-ground biomass to the soil surface. All biomass removed was bagged, dried, and weighed. Plot maintenance or removal of the target dominant species is performed annually or as needed.
Responses:
We estimate plant species cover for each live plant species within each 3 m x 4 m plot. Cover is estimated for all species as well as litter. Bare ground is calculated by subtracting the total cover of all species from 100%. Initial, pre-treatment measurements were made in 1996. Plant data are collected using standard SEV LTER protocols annually at peak fall biomass (Aug/Sept/Oct). Laboratory procedures: plant biomass removed from plots is dried, sorted by live and dead material, and weighed. Rain gauges were installed at each site and the corners of the areas containing each set of plots recorded in our ARCGIS database. Soil erosion bridges (1 m long) were installed in plots 1, 3 and 5 (removals and controls) at sites 1-5. We have recently measured soil biogeochemistry and texture.
Key Datasets:
Plant Removal Study: Recovery of Vegetation Following Disturbance at the Sevilleta National Wildlife Refuge, New Mexico. https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=168
Featured Publications:
Peters, D. P. C., and J. Yao. 2012. Long-term experimental loss of foundation species: consequences for dynamics at ecotones across heterogeneous landscapes. Ecosphere 3:art27. https://doi.org/10.1890/ES11-00273.1
Peters, D. P. C. 2002. Plant species dominance at a grassland–shrubland ecotone: an individual-based gap dynamics model of herbaceous and woody species. Ecological Modelling 152:5–32. https://doi.org/10.1016/S0304-3800(01)00460-4
Peters, D. P. C. 2002. Recruitment potential of two perennial grasses with different growth forms at a semiarid-arid transition zone. American Journal of Botany 89:1616–1623. https://doi.org/10.3732/ajb.89.10.1616

Mean-Variance Climate Experiment
Objective:
An innovative, cross-ecosystem experiment that factorially alters the mean and variance in soil moisture to simulate the effects of increasingly variable precipitation under a warming climate. This experiment would not be possible without long-term funding because it alters inter-annual variability.
Novelty:
The experiment has three innovations.
- determines interactions between reduced mean and increased variance for the first time, using a factorial design
- compares five dryland ecosystems to evaluate their susceptibility to transitions
- adds stochasticity to water manipulations to permit the antecedent effects that occur under natural climate variability
Design:
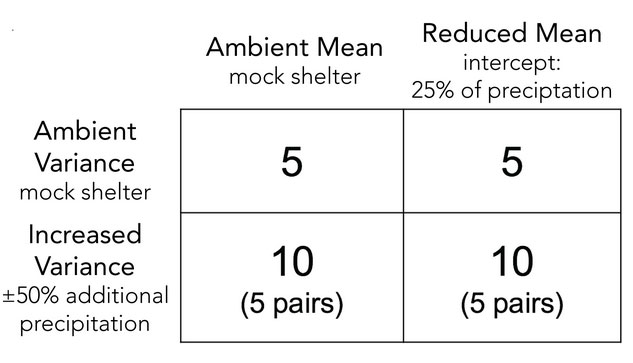
To increase variance in soil moisture (θ) stochastically, without changing the mean, we pair plots and amplify their precipitation regimes. Every water year, plots within a pair are randomly assigned to either a 50% decrease or 50% increase in precipitation. To reduce the long-run mean soil moisture (θ), we use roof panels to intercept 25% of precipitation year-round, a moderate forcing within the range of likely climate futures. Control plots receive similar shelters but with panels that allow precipitation throughfall. Plots receiving both reduced mean and increased variance randomly alternate between 75% less net precipitation (-25% for mean – 50% for variance) or 25% more (-25% for mean + 50% for variance). Replication is uneven to account for higher variability among plots in the increased variance treatment.










Water is captured from shelters with gutters, stored in tanks outfitted with a float switch, then delivered to the paired plots via solar-powered pumps in real time during rainfall events.
Responses:
Transition potential: Plant recruitment. Each year, we introduce seeds of “Current” and “Future” foundation plant species to test whether and at what rate a species introduced at low density are competitively excluded by resident dominants, a key determinant of species coexistence versus take-over. LTER core areas: We monitor above- and belowground plant production; plant demography via marked residents; biocrust cover and composition; decomposition; and plant, biocrust, and rhizospheric soil carbon and nitrogen. Sensors in 18 plots per ecosystem track hourly soil moisture and temperature at three depths (12.5, 22.5, 37.5 cm).
Supporting Documents:
Construction Documents | Collection Protocols
SEV-LTER Mean – Variance Experiment Plains Grassland Soil Moisture and Temperature. https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=334
SEV-LTER Mean – Variance Experiment Desert Grassland Soil Moisture and Temperature. https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=335

Monsoon Rainfall Manipulation Experiment
Objective:
The monsoon rainfall manipulation experiment (MRME) addresses the impacts of increased precipitation variability on the potential for plant community reordering in Chihuahuan Desert grassland by shrub encroachment. The overarching question is, How does the size and frequency of rain events affect post-fire ecosystem processes and the potential for shrub encroachment into grassland? MRME started in 2006 with Univ of New Mexico start-up funds to Collins, and is now supported by NSF LTREB and LTER.
Like most regions, models predict an increase in precipitation variability under climate change in the southwestern US. Although total monsoon precipitation has not changed over the past 100 years at SEV, the number of rain events per day increased slightly while average event size decreased slightly. In addition, wet years have only 2-3 additional large rain events relative to dry years. Where adults of black grama grass (Bouteloua eriopoda) and creosotebush (Larrea tridentata) co-occur, the fibrous-rooted black grama appears to be a better competitor for soil moisture than the more deeply rooted, but drought tolerant creosotebush. During establishment creosotebush seedlings must compete directly with black grama for shallow soil moisture. Thus, if changes to rain event size and frequency alter soil moisture depth and infiltration, then an increased frequency of small rain events could potentially increase the resistance of grassland to shrub invasion.
Novelty:
In comparison to experiments that manipulate the climate average (e.g., hotter, drier), many fewer studies have investigated how variability in climate affects populations, communities, and ecosystems. In our region, within-season variability is on the rise: The long-term pattern in climate data shows more frequent but smaller events have increased over the past century (Petrie et al. 2014). Our experiment detects the ecological consequences of changes in the regime, or pattern, of rainfall, while holding the total amount of rainfall constant. Over the long-term, we can newly evaluate the potential for interactions between within-season precipitation variability and interannual precipitation variability because MRME treatments occur over a highly variable background climate.
Design:
We manipulate rainfall event size and frequency using a fully randomized design with a total of thirteen 8 m x 13 m plots. Control plots (n=3) receive ambient precipitation. Two rainfall treatments (n=5) are applied each year during the monsoon. The Few-Large treatment receives ambient rainfall plus one 20 mm rain event each month (i.e., increased rainfall event size). The Many-Small treatment receives ambient rainfall plus four 5 mm rain events each month (i.e., increased rainfall event frequency). Thus, we add the same amount of rain over the monsoon season (60 mm), but we vary the size and frequency of applied precipitation events, increasing variability without imposing unrealistically extreme rainfall amounts. The overall precipitation added per monsoon season is equivalent to an increase of ~40% over an average year (long-term monsoon mean ~150 mm). The 20 mm events simulate more extreme precipitation events, and the 5 mm events represent modal precipitation event size. On average, 70% of natural precipitation events are <5 mm and 12% are between 10 – 20 mm (1988–2008). On August 4, 2009, a lightning-initiated fire began on the Sevilleta National Wildlife Refuge near MRME. The experimental site was entirely burned on this date, with all plots subjected to high intensity wildfire.











MRME research site, showcasing overhead sprinkler system, water tanks, water delivery, and “rainfall events” in action.
Responses:
MRME replicates are instrumented with soil moisture, temperature, and CO2 probes (Campbell Scientific CS107 temperature probes, Campbell Scientific CS-616 moisture probes buried at a 45o angle to obtain an integrated measure of moisture in the top 20 cm, Vaisala CARBOCAP CO2 sensors (GMM222, Vaisala, Helsinki, Finland) placed at three depths (2, 8, and 16 cm)). Each replicate has two 2 m x 2 m subplots, one of which is fertilized with 5 g N m-2 y-1 as NH4NO3. Aboveground net primary production (NPP) and plant species composition are measured in two 1-m2 quadrats in each fertilized and control subplot twice yearly. Belowground NPP is measured with root in-growth donuts.
Supporting Documents:
Mega plant files pending on EDI
Monsoon Rainfall Manipulation Experiment (MRME) Soil Temperature, Moisture and Carbon Dioxide Data from the Sevilleta National Wildlife Refuge, New Mexico. https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=304
Collins, S. 2020. Monsoon Rainfall Manipulation Experiment (MRME): Soil Nitrogen Data from the Sevilleta National Wildlife Refuge, New Mexico (2007 – 2020). https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=309
Monsoon Rainfall Manipulation Experiment (MRME): Net Primary Production Quadrat Data at the Sevilleta National Wildlife Refuge, New Mexico. https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=188
Collins, S. 2016. Monsoon Rainfall Manipulation Experiment (MRME) Meteorology Data from a Chihuahuan Desert Grassland at the Sevilleta National Wildlife Refuge, New Mexico (7/2007 – 8/2009). https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=188
Monsoon Rainfall Manipulation Experiment (MRME): Seasonal Biomass and Seasonal and Annual NPP Data at the Sevilleta National Wildlife Refuge, New Mexico. https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=206
Kwiecinski, J.V. 2019. Effects of Altered Precipitation on Biological Soil Crusts, Fungi, and Soil Nitrogen Availability at the Monsoon Rainfall Manipulation Experiment (MRME) in the Sevilleta National Wildlife Refuge, New Mexico (2016). https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=320
Related Publications:
Herrera, J., R. Poudel, K.A. Nebel and S.L. Collins. 2011. Precipitation increases the abundance of some groups of root-associated fungal endophytes in a semi-arid grassland. Ecosphere 2: https://doi.org/10.1890/ES11-00001.1
Thomey, M.L., S.L. Collins, R. Vargas, J.E. Johnson, R.F. Brown, D.O. Natvig, and M.T. Friggens. 2011. Effect of precipitation variability on net primary production and soil respiration in a Chihuahuan Desert grassland. Global Change Biology 17: 1505-1515. https://doi.org/10.1007/s00442-014-3052-1
Thomey, M. L., S. L. Collins, M. T. Friggens, R. F. Brown, and W. T. Pockman. 2014. Effects of monsoon precipitation variability on the physiological response of two dominant C-4 grasses across a semiarid ecotone. Oecologia 176:751–762. https://doi.org/10.1007/s00442-014-3052-1
Vargas, R., S.L. Collins, M.L. Thomey, J.E. Johnson, R.F. Brown, D.O. Natvig, and M.T. Friggens. 2012. Precipitation variability and fire influence the temporal dynamics of soil CO2 efflux in an arid grassland. Global Change Biology 18: 1401-1411. https://doi.org/10.1111/j.1365-2486.2011.02628.x
Kwiecinski, J. V., E. Stricker, R. L. Sinsabaugh, and S. L. Collins. 2020. Rainfall pulses increased short-term biocrust chlorophyll but not fungal abundance or N availability in a long-term dryland rainfall manipulation experiment. Soil Biology and Biochemistry 142:107693. https://doi.org/10.1016/j.soilbio.2019.107693
Petrie, M. D., S. L. Collins, D. S. Gutzler, and D. M. Moore. 2014. Regional trends and local variability in monsoon precipitation in the northern Chihuahuan Desert, USA. Journal of Arid Environments 103:63–70. https://doi.org/10.1016/j.jaridenv.2014.01.005

Nitrogen Fertilizer Experiment
Objective:
This long-term study examines how nitrogen fertilization affects plant species and soil microbial communities at the ecotone of Plains and Chihuahuan Desert grassland. The project at the Sevilleta LTER began in spring 1995 with funding from an NSF special competition for cross-site research. LTER funds now maintain treatments and data collection.
Novelty:
This is one of the longest running dryland nitrogen fertilization experiments, contributing to our general understanding of the effects of terrestrial eutrophication, and providing an important dryland representative for cross-site studies of N limitation in grasslands and other plant communities. Over the long-term, we can evaluate the potential for interactions between eutrophication and increases in both aridity and interannual precipitation variability because fertilization treatments occur over a highly variable and drying background climate. For example, nitrogen addition may alleviate a saturating relationship between plant biomass and precipitation and reduce the sensitivity of biomass to variability in climate.
Design:
Fertilizer is applied yearly prior to the summer monsoon rainy season as granular NH4NO3 at the rate of 100 kg N ha-1 yr-1. The twenty 10 m X 5 m plots include 10 fertilized and 10 control plots. This site was burned in a management fire in June 2003. A management fire in September 2019 burned around, but not in, this experiment.
Responses:
Plant species composition and the cover and height of individuals to estimate biomass, were sampled twice yearly (spring and fall) within four permanent 1 m x 1 m quadrats until 2019, and afterwards two quadrats per plot. Belowground plant biomass (NPP) is measured in root in-growth donuts annually.
Supporting Documents:
Nitrogen Fertilization Experiment (NFert): Seasonal Biomass and Seasonal and Annual NPP Data at the Sevilleta National Wildlife Refuge, New Mexico. https://portal.edirepository.org/nis/mapbrowse?scope=knb-lter-sev&identifier=186
Related Publications:
Zeglin, L.H., M. Stursova, R.L. Sinsabaugh & S.L. Collins. 2007. Microbial responses to nitrogen addition in three contrasting grassland ecosystems. Oecologia 154:349-359. https://doi.org/10.1007/s00442-007-0836-6
Stursova, M., C.L. Crenshaw & R.L. Sinsabaugh. 2006. Microbial responses to long-term N deposition in a semi-arid grassland. Microbial Ecology 51:90-98. https://doi.org/10.1007/s00248-005-5156-y
Ladwig, L.M., S.L. Collins, A.L. Swann, Y. Xia, M.F. Allen and E.B. Allen. 2012. Above- and belowground responses to nitrogen addition in a Chihuahuan Desert grassland. Oecologia 169: 177-185. https://doi.org/10.1007/s00442-011-2173-z
Porras-Alfaro, A., J. Herrera, R.L. Sinsabaugh, K.J. Odenbauch, T. Lowrey & D.O. Natvig. 2008. Novel root fungal consortium associated with a dominant desert grass. Applied and Environmental Microbiology 74:2805-2813. https://doi.org/10.1128/AEM.02769-07
Porras-Alfaro, A., J. Herrera, D. O. Natvig, and R. L. Sinsabaugh. 2007. Effect of long-term nitrogen fertilization on mycorrhizal fungi associated with a dominant grass in a semiarid grassland. Plant and Soil 296:65–75. https://doi.org/10.1007/s11104-007-9290-9